SCI论文(www.lunwensci.com):
摘要:目的构建以miRNA为中心的综合调控网络,探讨关键miRNA、靶基因、转录因子、长链非编码RNA与先天性小耳畸形发病间的关系,为进一步明确病因提供新的研究思路。方法分析小耳畸形残耳软骨与健侧耳软骨间miRNAs差异表达谱,基于24个差异表达的miRNAs,预测靶基因、转录因子和长链非编码RNA,构建综合调控图并对关键节点进行生物信息学分析。
结果基于24个差异miRNAs,共预测到154个靶基因,209个转录因子和22个长链非编码RNA。MiR-221/222-3p、miR-320c、miR-3122、miR-4521,靶基因ZEB2、TRPS1、GNAI3、SHOC2、PDPK1、PDPK1,转录因子EP300、STAT1、FOS、KMT2A、RNF2和长链非编码RNA TUG1、MIAT间存在相互调节作用构成关键模块。MiR-221/222-3p可与靶基因TRPS1、ZEB2,转录因子FOS、EP300、STAT1及lncRNA MIAT相互调控,靶基因ZEB2与转录因子STAT1在调控网络中作为关键节点可连接多个靶点构成综合调控网络。
结论初步建立了先天性小耳畸形综合调控网络,找到了表达异常的关键miRNAs、靶基因、转录因子和长链非编码RNA,为后期针对各个靶点的验证和基因功能的研究奠定了理论基础,利于进一步探讨关键靶点及其调控网络与先天性小耳畸形间的关系。
关键词:先天性小耳畸形;microRNA;靶基因;长链非编码RNA;调控网络
本文引用格式:郭蕊,杨庆华.先天性小耳畸形微小RNA综合调控网络的生物信息学分析[J].世界最新医学信息文摘,2019,19(51):114-117.
0引言
先天性小耳畸形(Congenital Microtia)是外耳和中耳的先天发育畸形,由胚胎时期第一鳃沟及其附近第一、二鳃弓发育异常引起,常表现为耳廓畸形、外耳道闭锁和(或)中耳畸形,并伴有不同程度听力损害。先天性小耳畸形发病率为0.83-17.40/万[1],我国约3.06/万,且有逐年上升趋势[2]。
目前,由于染色体畸变和单基因缺陷等,已发现有超过18种不同的与小耳畸形相关的综合征,因此,遗传因素发挥着重要作用[1]。研究发现微小RNA(microRNA,miRNA)参与小耳畸形的发病,但对其在先天性小耳畸形中发挥的具体作用和机制尚不清楚[3]。MiRNA是一类20-24个核苷酸的非编码RNA,通过与靶mRNA的3'UTR区域的互补结合来调节翻译水平的基因表达,调节细胞的生长、分化和凋亡[4]。
探讨miRNA在先天性小耳畸形中的作用,构建miRNAs-靶基因-长链非编码RNA(Long non-coding RNA,LncRNA)-转录因子(Transcription factor,TF)综合调控网络,将从一个全新的角度诠释和理解先天性小耳畸形的形成机制,为今后的研究及临床工作提供新的指导方向。
1材料与方法
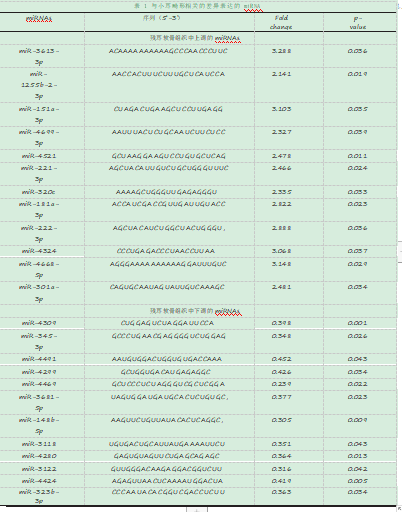
1.1数据来源。宋[3]等人将先天性小耳畸形患者的残耳软骨组织作为研究对象(患者均为最常见的II°畸形),以同一患者的健侧正常耳软骨作为自身对照,通过对实验组3例及对照组3例样本的基因组miRNAs进行扫描,筛选出24个差异表达的miRNAs(筛选标准:Fold change≥2,P<0.05),其中实验组相对于对照组上调12个,下调12个(表1)。
1.2靶基因预测。利用网络资源预测靶基因并筛选出所用平台均可预测到的结果。预测平台包括:miRDB(http://www.mirdb.org/)、TargetScan(http://www.targetscan.org/)、miRWalk(http://mirwalk.umm.uni-heidelberg.de/)RNA22(http://cbcsrv.watson.ibm.com/rna22.html)miRTarbase(http://mirtarbase.mbc.nctu.edu.tw/php/index.php)[5]。
1.3MiRNA与lncRNA间调控关系的预测。
StarBase v3.0(http://starbase.sysu.edu.cn/index.php数据库可用于系统地识别RNA-RNA和RNA-蛋白质间的相互作用[5]。我们从starBase v3.0中下载了所有miRNA-lncRNA交互网络,并筛选出与24个miRNAs相关的所有子网络。
1.4TFs与miRNA间调控关系预测。TransmiR v2.0数据库是有关TFs-miRNA调控的数据库[5],此次研究中,我们下载了有关24个miRNAs的TF-miRNA子网络以便进一步分析。
2结果
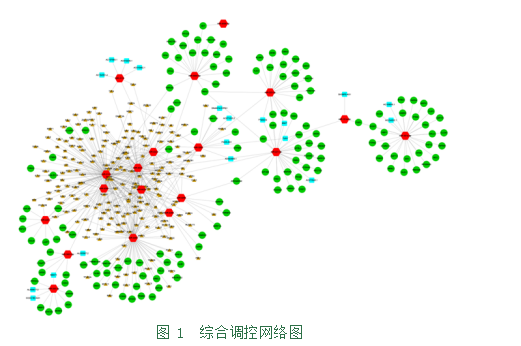
2.1构建综合调控网络。综合以上结果,构建包含miRNA-靶基因、TF-miRNA和miRNA-LncRNA关系的调控图(图1),发现miR-221-3p、miR-222-3p、miR-4668-5p、miR-301a-3p、miR-4469可调节一系列靶基因;miR-221-3p、miR-320c、miR-345-3p、miR-3118可调节一系列LncRNAs;miR-4521、miR-3122可被一系列TFs调节。其中红色六边形代表 miRNA,绿色圆圈代表靶基因,橙色三角形代表 TF,蓝色正方形代表 lncRNA。
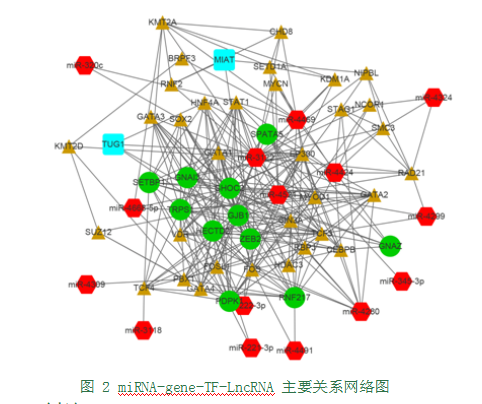
筛选关键调控网络。通过文献查询及预测结果,筛选出与胚胎发育、耳发育、软骨发育、先天性疾病等相关的靶点, 初步得到miRNA-gene-LncRNA-TFs 主要的综合调控图(图2)。
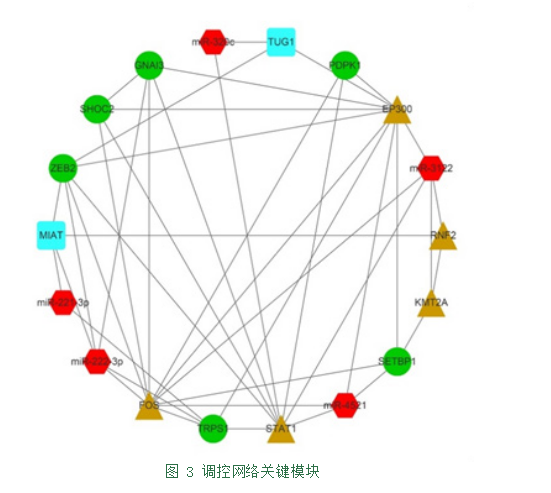
再根据各个节点间连接度的大小,筛选出关键模块(图3)。
3讨论
本研究从 miRNA 角度对先天性小耳畸形进行研究,致力于寻找新的致病靶点及调控途径。通过生物信息学分析构建调控网络,发现miR-221/222-3p、miR-320c、miR-3122、miR-4521,靶基因ZEB2、TRPS1、GNAI3、SHOC2、PDPK1、PDPK1,转录因子EP300、STAT1、FOS、KMT2A、RNF2,长链非编码RNA TUG1、
MIAT在先天性小耳畸形中可能起关键作用。
生物信息学分析发现图3中,miR-221/222-3p、TRPS1、ZEB2、FOS、EP300、STAT1及lncRNA TUG1可
构成第一个调控网络。MiR-221/222-3p编码基因位于X染色体p11.3附近,二者属同一家族在X染色体上同步表达。在乳腺癌中,miR-221/222介导的E-钙粘蛋白丰度的降低依赖于TRPS1的3’非翻译区,而TRPS1通过降低ZEB2基因的表达可抑制乳腺癌细胞上皮-间质转化[6]。
研究发现miR-221-3p在人软骨组织中表达下调且其表达水平与软骨退化程度有关,其表达上调可阻止软骨细胞外基质(Extracellular Matrix,ECM)降解,也可通过调节基质金属蛋白酶-13(matrix metalloproteinase 13,MMP13)影响ECM,影响软骨细胞表型转变[7]。
TRPS1作为转录抑制因子,可特异结合GATA序列,抑制脊椎动物发育中特定阶段和位点的GATA调节基因的表达,调节软骨细胞增殖和分化。此基因突变可导致毛发-鼻-指(趾)综合征(Tricho-rhino-phalangeal syndrome,TRPS),此类患者身材矮小,髋关节异常,锥形骨骺和生长板过早闭合,反映出软骨内骨化缺陷,且动物实验发现TRPS的骨骼发育不良主要是软骨细胞和软骨膜发育失调引起[8]。
ZEB2基因编码蛋白质可在神经嵴衍生细胞和中枢神经系统模式内及人类中线结构的发育中起作用[9]。动物实验发现敲除ZEB2基因的小鼠出现颅神经嵴细胞(Cranial neural crest cell,CNCCs)的分层停滞,且敲除ZEB2的胚胎在其第9.5天时死亡,发现其神经管闭合失败,神经板和外胚层其余部分之间缺乏清晰的边界且存在第一鳃弓缺失,表明ZEB2基因产物对迷走神经嵴前体的发育和小鼠CNCCs的迁移必不可少[10]。FOS基因编码亮氨酸拉链蛋白,与JUN家族的蛋白质构成转录因子复合物AP-1[11]。
在三阴性乳腺癌中,AP-1可通过上调ZEB2转录,抑制E-钙粘蛋白表达并刺激细胞侵袭[12]。且Errico[13]等人证明了c-FOS是miR-221/222的直接靶点,且确定了HOXB7/PBX2→miR-221/222→c-FOS调节链,而HOXB7/PBX2二聚体的功能性丧失可致miR-221/222转录降低及c-FOS表达升高,致细胞死亡。EP300基因编码组蛋白乙酰转移酶,可通过染色质重塑调节转录并在细胞增殖和分化过程中起重要作用。
EP300的靶点之一是FOS基因,其可能在转录书签或预处理过程中影响细胞的生长发育[14]。EP300也可激活ZEB2启动子,使进展期乳腺癌获得侵袭性表型[15]。STAT1即信号转导和转录因子1,其参与的JAK/STAT信号通路是细胞因子受体下游的广泛信号传导途径[16]。动物实验发现STAT1可介导胰岛素样生长因子1在大鼠软骨细胞中起独立凋亡作用[17],STAT1不仅可以直接调节细胞周期,还可调节成纤维细胞生长因子受体(fibroblast growth factor receptor,FGFR)的表达[18]。
Ahmad[19]等人发现激活STAT1与CBP/P300复合物可抑制软骨细胞中MMP13基因的转录诱导。通过KEGG通路分析,发现在JAK/STAT信号通路内,STAT1可激活CBP/P300介导信号传导和基因表达。研究发现LncRNA TUG1在膀胱肿瘤中表达下调可通过靶向ZEB2并使Wnt/β-catenin信号通路失活,抑制肿瘤增殖并诱导其凋亡[20]。
其次,生物信息学分析发现图3中,SETBP1、KMT2A、RNF2、ZEB2及lncRNA TUG1可构成第二个调控网络。其中SETBP1可通过募集HCF1/KMT2A/PHF8表观遗传复合物引起基因表达的激活[21]。KMT2A又称混合谱系白血病(Mixed Lineage Leukemia,MLL)基因,MLL编码蛋白可与HOXA9连接,与PcG(polycomb group,PcG)蛋白相互作用调节HOX基因[22]。RNF2也可与PcG蛋白形成多蛋白复合物,具有泛素连接酶(E3)活性,参与发育与细胞增殖等各种调节基因的转录抑制[23]。
动物实验发现RNF2同源基因Ring1b纯合小鼠具有轴向骨骼异常并在中胚层隔室正常边界前的细胞中抑制HOX基因表达。Ring1b在鸡胚中的过表达可抑制神经管中HOXB9表达,且敲除RNF2基因的小鼠表现出原肠胚形成停滞。由此,Suzuki、Voncken等人认为RNF2参与小鼠前-后体轴的发育以及早期哺乳动物发育中的细胞增殖[24-25]。而LncRNA TUG1可通过与PcG复合物2的相互作用抑制基因表达[23]。
最后,SHOC2、PDPK1与STAT1可构成第三个调控网络。SHOC2作为蛋白磷酸酶1(PP1c)的调节亚基,充当M-Ras/MRAS的效应器并参与MAPK激活途径。Selfors[26]等发现SHOC2基因编码的蛋白质参与了FGFR信号转导。PDPK1即3-磷酸肌醇依赖性蛋白激酶1,敲除PDPK1基因的小鼠可出现体节、前脑和神经嵴衍生组织的缺失[27],其也可通过p38 MAPK信号通路促进骨关节炎中软骨细胞的凋亡[28]。
通过以上分析,我们发现miR-221/222-3p、TRPS1、ZEB2、FOS、EP300、STAT1及lncRNA TUG1构成的第一个调控网络主要作用于软骨细胞及ECM等,而耳廓软骨发育异常正是小耳畸形最突出、最多样化的特征。SETBP1、KMT2A、RNF2、ZEB2及lncRNA TUG1构成第二个调控网络可能通过与PcG复合物相互作用影响CNCCs及同源异形盒基因HOX的调控。HOX基因可调节胚胎时期第二鳃弓内CNCCs前-后体轴的发育,只有携带耳廓各个部分正常发育信息的CNCCs在正确时间内迁入正确区域,才能形成正常耳廓,一旦其迁移路径、时间或区域受到干扰,则会形成畸形耳廓[1]。SHOC2、PDPK1与STAT1构成的第三个调控网络主要涉及MAPK及FGF信号通路。
FGF是CNCCs的关键成活因子,其参与的信号通路已证明与小耳畸形的发生有关[1]。进一步分析发现,ZEB2及STAT1作为关键节点可将三个调控网络相连。鉴于FOS、EP300及STAT1均可参与JAK/STAT信号通路作用于MMP13,而miR-221/222-3p也可作用于MMP13影响ECM,由此推论miR-221/222-3p可能参与JAK/STAT信号通路发挥调控作用。
目前,虽尚未有文献报道lncRNA MIAT与软骨发育、胚胎发育、器官形成等先天性小耳畸形发生因素相关,但图3中,MIAT与miR-221/222-3p、ZEB2、RNF2相关连,由此推断MIAT可能同上述基因相互调节,通过影响CNNCs或HOX表达影响软骨发育,参与小耳畸形的发生发展。同理,miR-3122在图3中与STAT1、RNF2、KMT2A、EP300及FOS相关,推测其可能作为关键节点与基因ZEB2通过调节胚胎发育,共同参与第一、二调控网络。而PDPK1与转录因子FOS、EP300、STAT1相关,推测PDPK1可能参与JAK/STAT信号通路。
此次研究初步建立了先天性小耳畸形的综合调控网络,找到了关键miRNAs、靶基因、转录因子和长链非编码RNA。利于进一步探讨关键靶点及其调控网络与先天性小耳畸形发生发展之间关系,为后期针对各靶点的验证和基因功能的相关研究奠定了理论基础。
但此次研究仍存在一些局限性,如在数据分析过程中,样本量不足可能导致结果假阳性率较高,因此,还需进一步通过RT-PCR、Western blot及动物实验等进行遗传和实验室验证;在生物信息学分析过程中,一部分研究较少的基因可能存在信息不准确,导致在检索过程中产生误差等。总之,我们的研究数据提供了可能参与先天性小耳畸形发生的与miRNA相关靶点的全面的生物信息学分析,为以后的科研及临床工作奠定理论基础。
参考文献
[1]Luquetti D V,Heike C L,Hing A V,et al.Microtia:epidemiolo gy and genetic s[J].Am J Med G enet A,2012,158A(1):124-139.
[2]Deng K,Dai L,Yi L,et al.Epidemiologic characteristics and time trend in the prevalence of anotia and microtia in China[J].Birth Defects Res A Clin Mol Teratol,2016,106(2):88-94.
[3]宋宇鹏,杨庆华,林琳,等.先天性小耳畸形microRNA及mRNA表达谱的变化及关键靶基因的筛选[J].医学研究杂志,2017(06):109-113.
[4]Shruti K,Shrey K,Vibha R.Micro RNAs:tiny sequences with enormous potential[J].Biochem Biophys Res Commun,2011,407(3):445-449.
[5]Shukla V,Varghese V K,Kabekkodu S P,et al.A compilation of Web-based research tools for miRNA analysis[J].Brief Funct Genomics,2017,16(5):249-273.
[6]Stinson S,Lackner M R,Adai A T,et al.TRPS1 targeting by miR-221/222 promotes the epithelial-to-mesenchymal transition in breast cancer[J].Sci Signal,2011,4(177):a41.
[7]Zheng X,Zhao F C,Pang Y,et al.Downregulation of miR-221-3p contributes to IL-1beta-induced cartilage degradation by directly targeting the SDF1/CXCR4 signaling pathway[J].J Mol Med(Berl),2017,95(6):615-627.
[8] Napierala D,Sam K,Morello R,et al.Uncoupling of chondrocyte differentiation and perichondrial mineralization underlies the skeletal dysplasia in tricho-rhino-phalangeal syndrome[J].Hum Mol Genet,2008,17(14):2244-2254.
[9]Amiel J,Espinosa-Parrilla Y,Steffann J,et al.Large-scale deletions and SMADIP1 truncating mutations in syndromic Hirschsprung disease with involvement of midline structures[J].Am J Hum Genet,2001,69(6):1370-1377.
[10]Van de Putte T,Maruhashi M,Francis A,et al.Mice lacking ZFHX1B,the gene that codes for Smad-interacting protein-1,reveal a role for multiple neural crest cell defects in the etiology of Hirschsprung disease-mental retardation syndrome[J].Am J Hum Genet,2003,72(2):465-470.
[11]Wang Z,Xu M,Bai J,et al.Vildagliptin reduced extracellular matrix degradation in human primary chondrocytes[J].Eur J Pharmacol,2019(844):49-55.
[12]Zhao C,Qiao Y,Jonsson P,et al.Genome-wide profiling of AP-1-regulated transcription provides insights into the invasiveness of triple-negative breast cancer[J].Cancer Res,2014,74(14):3983-3994.
[13]Errico M C,Felicetti F,Bottero L,et al.The abrogation of the HOXB7/PBX2 complex induces apoptosis in melanoma through the miR-221&222-c-FOS pathway[J].Int J Cancer,2013,133(4):879-892.
[14]Byun J S,Wong M M,Cui W,et al.Dynamic bookmarking of primary response genes by p300 and RNA polymerase II
complexes[J].Proc Natl Acad Sci U S A,2009,106(46):19286-
19291.
[15]Cho M H,Park J H,Choi H J,et al.DOT1L cooperates with the c-Myc-p 300 complex to epigenetically derepress CDH1 transcription factors in breast cancer progression[J].Nat Commun,2015(6):7821.
[16]Rosenzweig S D,Holland S M.Defects in the interferon-gamma and interleukin-12 pathways[J].Immunol Rev,2005,(203):38-47.
[17]Spagnoli A,Torello M,Nagalla S R,et al.Identification of STAT-1 as a molecular target of IGFBP-3 in the process of chondrogenesis[J].J Biol Chem,2002,277(21):18860-18867.
[18]Xiao L,Naganawa T,Obugunde E,et al.Stat1 controls postnatal bone formation by regulating fibroblast growth factor signaling in osteoblasts[J].J Biol Chem,2004,279(26):27743-27752.
[19]Ahmad R,Qureshi H Y,El M M,et al.Inhibition of interleukin 1-induced matrix metalloproteinase 13 expression in human chondrocytes by interferon gamma[J].Ann Rheum Dis,2007,66(6):782-789.
[20]Liu Q,Liu H,Cheng H,et al.Downregulation of long noncoding RNA TUG1 inhibits proliferation and induces apoptosis through the TUG1/miR-142/ZEB2 axis in bladder cancer cells[J].Onco Targets Ther,2017(10):2461-2471.
[21]Piazza R,Magistroni V,Redaelli S,et al.SETBP1 induces transcription of a network of development genes by acting as an epigenetic hub[J].Nat Commun,2018,9(1):2192.
[22]]Pradeepa M M,Grimes G R,Taylor G C,et al.Psip1/Ledgf p75 restrains Hox gene expression by recruiting both trithorax and polycomb group proteins[J].Nucleic Acids Res,2014,42(14):9021-9032.
[23]Khalil A M,Guttman M,Huarte M,et al.Many human large intergenic noncoding RNAs associate with chromatin-modifying complexes and affect gene expression[J].Proc Natl Acad Sci U S A,2009,106(28):11667-11672.
[24]Suzuki M,Mizutani-Koseki Y,Fujimura Y,et al.Involvement of the Polycomb-group gene Ring1B in the specification of the anterior-posterior axis in mice[J].Development,2002,129(18):4171-4183.
[25]Voncken J W,Roelen B A,Roefs M,et al.Rnf2(Ring1b)deficiency causes gastrulation arrest and cell cycle inhibition[J].Proc Natl Acad Sci U S A,2003,100(5):2468-2473.
[26]Selfors L M,Schutzman J L,Borland C Z,et al.soc-
2 encodes a leucine-rich repeat protein implicated in fibroblast growth factor receptor signaling[J].Proc Natl Acad Sci U S A,1998,95(12):6903-6908.
[27]Lawlor M A,Mora A,Ashby P R,et al.Essential role of PDK1 in regulating cell size and development in mice[J].EMBO J,2002,21(14):3728-3738.
[28]Ge Q,Wang H,Xu X,et al.PDK1 promotes apoptosis of chondrocytes via modulating MAPK pathway in osteoarthritis[J].Tissue Cell,2017,49(6):719-725.
关注SCI论文创作发表,寻求SCI论文修改润色、SCI论文代发表等服务支撑,请锁定SCI论文网!
文章出自SCI论文网转载请注明出处:https://www.lunwensci.com/yixuelunwen/11718.html