SCI论文(www.lunwensci.com):
摘要:目的血小板活化因子(Platelet-activating factor,PAF)促进精子受精潜能,从而提高受精率和临床妊娠率。方法通过FITC-PSA染色确定顶体状态、酪氨酸磷酸化检测、细胞外信号调节激酶活性(ERK)的检测来分别分析PAF对精子顶体反应、精子蛋白酪氨酸磷酸化及人精子ERK信号通路的作用。结果PAF处理显著(P<0.05)提高了精子顶体反应的百分率;不同浓度PAF孵育对人精子的运动无影响,所有浓度的PAF处理均能诱导精子蛋白磷酸化酪氨酸的表达;在浓度为10nM的PAF处理下,人精子中ERK1和ERK2蛋白水平显著升高;使用浓度>0.1μM的U0126预处理可显著并且剂量依赖性地抑制了PAF诱导的人精子顶体反应(P<0.05),U0124对PAF诱导的人精子顶体反应无影响。结论PAF诱导精子顶体反应和酪氨酸磷酸化。PAF通过促进ERK1和ERK2的酶活性激活人精子中的ERK信号通路介导对精子顶体反应的提高。我们的研究结果进一步验证了PAF对人类精子受精能力的潜在作用。
关键词:血小板活化因子;顶体反应;酪氨酸磷酸化;人精子ERK信号通路;人类精子受精能力
本文引用格式:吴海涛,黎平,陈希曦,等.血小板活化因子对人精子生物学活性和细胞外信号调节激酶的影响[J].世界最新医学信息文摘,2019,19(59):159-162.
0引言
顶体反应是由顶体膜与质膜融合、水解酶释放和透明带降解构成的一个多步骤的过程,是精子穿透透明带的关键。由于顶体反应对受精至关重要,其通常被认为是精子受精能力的间接评价指标。
血小板活化因子(1-O-alkyl-2-O-acetyl-sn-glycero-3-phosphorylcholine,PAF)是一种高效的磷脂激活剂和多种生物活性介质[1-3]。其非生物活性和去乙酰化的前体为1-alkyl-2-lyso-sn-glycero-3-phosphocholine(Lyso-PAF)[4]。PAF存在于人类精子中,其在男性生殖系统中的作用已被广泛研究。研究已显示精子内源性PAF的浓度与精子活力和发育有关[5-7]。而外源性PAF可通过增加ATP含量促进精子受精潜能[8]。虽然PAF明显参与调节精子的受精能力,但其调控精子参数的潜在机制尚未明确。
细胞外信号调节激酶(ERK)属于丝裂原活化蛋白激酶(MAPK)家族。它们是丝氨酸/苏氨酸激酶,即需要对苏氨酸和酪氨酸残基进行双重磷酸化才能完全激活[9]。已知ERK信号通路在哺乳动物精子中表达,参与精子活力,获能和顶体反应的激活[10-13]。在这项研究中,我们假设PAF通过激活ERK信号通路调节人类精子的顶体反应和运动。
1材料和方法
1.1精液收集和精子准备
本研究已获香港大学伦理委员会小组批准。精液样本取自就诊香港玛丽医院亚生育科诊所的男性。收集前须先取得书面同意书。研究使用的精液均符合WHO精液参数正常标准(WHO,1998)。
精液液化完全后,采用上游法于精液中分离精子(WHO,1998)。简而言之,将1.2 mL EBSS/0.3%BSA轻轻置于1 mL精液样本之上,置于15 mL无菌锥形离心管中。试管斜置45°,于37℃,5%CO2孵育1小时。然后轻轻地回到直立的位置。仔细收集最上层1 mL EBSS/0.3%BSA,300g离心10分钟。收集到的球团经过两次洗涤后,重悬于EBSS/3%BSA过夜放置以获能。除非另有说明,本研究使用的所有样本均为获能精子。实验当天,取500g离心5分钟,重悬于EBSS/0.3%BSA中。
1.2通过FITC-PSA染色确定顶体状态
如前文所述进行荧光素异硫氰酸酯标记的花生(Pisum sativum)凝集素(FITC-PSA)(Sigma,U.S.A.)和Hoechst染色[14]。将处理过的精子样品在5l磷酸盐缓冲盐水(PBS)含有0.05gsupravital染色剂Hoechst 33258(双苯甲亚胺;Sigma)构成的染色溶液中温育15分钟。其后,在1 mL 2%(w/v)聚乙烯吡咯烷酮-40的PBS(pH7.4)溶液中以900g离心10分钟洗去样品中染料。重悬精子沉淀,并在300l95%乙醇中4℃放置30分钟。使用95%乙醇将最终精子浓度调节至10-20×106个/mL。将精子涂在载玻片上,室温风干。随后将载玻片用含有2gFITC-PSA的染色溶液在20lPBS中染色30分钟,蒸馏水洗涤两次并固定在含有0.2M正丙基丙酸酯(Sigma,U.S.A.)的甘油中。
在400倍放大的荧光显微镜下对至少200个随机选择的精子进行定量。使用精子活力和顶体状态对精子进行评分。由于Hoechst染色在与DNA结合时表现出强烈的蓝色荧光信号,故具有强Hoechst染色(Hoechst阳性)的精子被认定为死亡精子。FITC-PSA则作为顶体外膜存在的标志。根据凝集素染色模式可对顶体状态进行分类:(i)完整顶体:顶体的完全染色;(ii)反应中顶体:顶体部分或斑片状染色,无Hoechst染色;(iii)反应后顶体:赤道区染色或整个精子头未见染色,无Hoechst染色。
使用cell-UV一次性精液分析室(Fertility Technologies,Inc.,Natick,MA)结合计算机辅助精子分析(CASA)系统(美国Hobson精子追踪系统)分析精子活力。每次测量均在37℃的加热显微镜下进行,10分钟内完成。设置的参数系统有:(i)成帧率:60 Hz,(ii)纵横比:1.39,(iii)校准:10.0,(iv)刷新时间:5s,(v)阈值+20/-20,(vi)滤波器重量:1,-1;2,0;3,0;4,0,(vii)最小室深度:20m,(viii)最小轨迹点:25,(ix)最大轨迹点:250,(x)轨迹绘制时间:3秒。对于每次评估,对来自随机选择的区域的200个精子进行定量。CASA系统根据以下参数测量精子活力:
(1)曲线速度(VCL;m/s)-精子头沿曲线路径移动的平均速度。
(2)直线速度(VSL;m/s)-精子头在其第一个和最后一个检测位置之间线性移动的平均速度。
(3)线性度(LIN;%)-沿曲线路径的线性运动的度量,表示为VSL/VCL。
(4)鞭打频率(BCF;Hz)-曲线路径与平均路径相交的平均速率。
(5)头侧摆幅度(ALH;m)-精子头部偏离其平均路径轴线的幅度。
1.3酪氨酸磷酸化检测
取浓度为2×106/mL的精子,600 g离心10分钟,1.5 mL TBS冲洗一次。精子团重悬于裂解缓冲液(Cytobuster,Novagen),加入蛋白酶和磷酸酶抑制剂混合物(Calbiochem),室温孵育1小时。样品以20,000g离心10分钟后,仔细收集上清,煮沸5分钟后,使用10%凝胶在Mini-PROTEIN 3 Electrophoresis System(BioRad)中进行SBS-PAGE电泳。凝胶在恒压115 V的聚氯乙烯氟化膜(Millipore,Bedford,MA)上印迹75分钟。然后使用专为酪氨酸磷酸化设计的单克隆抗体(PY20,Sigma)进行Western blot分析,随后使用与碱性磷酸酶(Novagen)结合的山羊抗小鼠IgG进行免疫印迹分析。采用多克隆抗微管蛋白抗体(Sigma)检测各泳道的加载样本。
1.4细胞外信号调节激酶活性(ERK)的检测
实验处理后的精子(10×106个/mL)使用PBS冲洗三次。精子蛋白在50l的蛋白提取缓冲液(CentobusterTM,Calbiochem,CA,USA)中用蛋白酶抑制剂混合物(Calbiochem)在室温下提取1小时。20 000×g离心10分钟后收集上清液,随后用12%SDS-PAGE对精子溶解液进行分解,并在PVDF膜上进行印迹。采用单克隆抗活化ERK抗体(克隆MAPK-YT,0.5g/mL;sigma)进行Western blot分析,随后用山羊抗小鼠二级抗体结合辣根过氧化物酶进行检测。采用抗微管蛋白抗体(Sigma)检测每个泳道的加载样本。非放射性ERK1/2活性测定试剂盒(Chemicon,Temecula,usa)也被用来根据制造商的说明确认Western blot的结果。结果以未治疗组与对照组比较的百分比变化表示。
1.5统计分析
结果用均数±SEM(均数标准误差)表示。数据均采用统计软件进行分析(SigmaPlot 8.02&SigmaStat 2.03,Jandel Scientific,San Rafael,USA)。所有实验均采用多重比较秩检验的非参数方差分析。在适当的情况下,使用参数学生t检验或非参数Mann Whitney U检验作为后检验。P<0.05认为差异有统计学意义。
2结果
2.1PAF增强人精子顶体反应
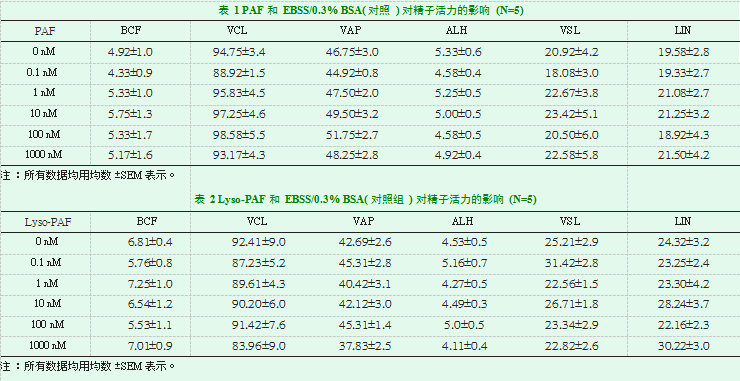
如图1A所示,PAF处理(0.1-1000nM)显著(P<0.05)提高了精子顶体反应的百分率,且呈剂量依赖性(N=5)。而不同浓度(0.1-1000nM)的Lyso-PAF对精子顶体反应无影响(图1B)。
2.2PAF对人精子的运动无影响。
如表1所示,在不同浓度(0.1-1000nM)的PAF下孵育90分钟后,精子活动性的参数(包括精子BCF,VCL,VAP,VSL,LIN)均无显著变化。同样,在所有测试浓度(0.1-1000nM)的Lyso-PAF下孵育90分钟后,人精子活动性参数也无显著变化(表2)。
2.3PAF诱导精子蛋白酪氨酸磷酸化
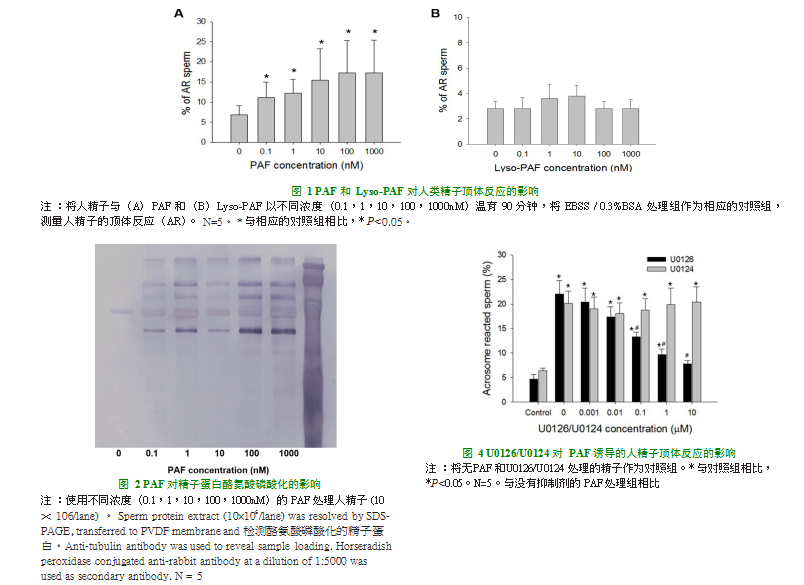
所有浓度(0.1,1,10,100,1000nM)的PAF处理均能诱导精子蛋白磷酸化酪氨酸的表达(图2)。
2.4PAF激活人精子ERK信号通路
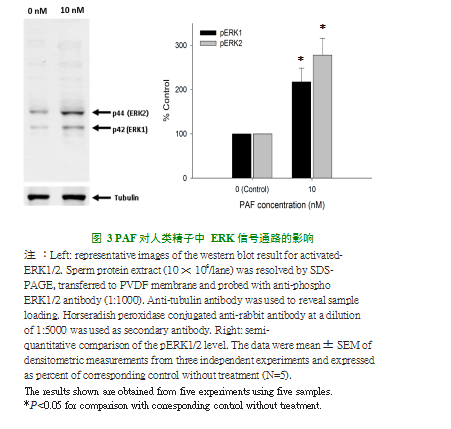
如图3所示,在浓度为10nM的PAF处理下,人精子中ERK1和ERK2蛋白水平显著升高。这表明,PAF处理能激活人精子ERK信号通路。
2.5ERK途径抑制剂对透明带诱导顶体反应的影响
在存在和不存在ERK途径抑制剂U0126和U0124的情况下,比较PAF诱导的顶体反应的获能精子百分比(图4)。使用浓度>0.1M的U0126预处理可显著并且剂量依赖性地抑制了PAF诱导的人精子顶体反应(P<0.05)。另一方面,U0124对PAF诱导的人精子顶体反应无影响(图4)。
3讨论
PAF是一种乙酰化甘油磷脂,在许多细胞类型中具有多种生物学功能[3]。其参与调节如白细胞功能、血小板聚集和脱颗粒以及炎症等生物活性[15],在排卵、卵母细胞受精、胚胎早期发育、着床、分娩等生殖过程中的作用也得到了广泛的研究。此外,PAF在调节精子功能方面发挥着至关重要的作用。内源性PAF浓度与人精子受精潜能及妊娠结局相关[18]。近期的研究表明,采用外源性PAF处理的精子可显著提高体内妊娠率,并增强精子在体外对无透明带仓鼠卵的穿透能力[19]。此外,用PAF处理的精子受精的卵母细胞可改善囊胚的形成和发育[20]。
本研究的数据表明PAF对人类精子顶体反应率具有剂量依赖性影响。PAF处理精子90分钟可诱导酪氨酸磷酸化并激活ERK信号通路,对精子运动的影响最小,并且ERK抑制剂U0126可显着阻断PAF诱导的顶体反应。然而,作为PAF前体的Lyso-PAF对顶体反应和精子活力无明显影响。
我们的研究结果与既往的发现一致,即PAF可促进人类精子的获能和顶体反应[21]。在一项研究的26份样本中,有20份在接受10 nM PAF治疗后,顶体反应率显著升高,并可被PAF拮抗剂抑制[22]。此外,有研究显示人精子中PAF浓度与精子穿透能力呈正相关,反之亦然[18]。也有报道称,PAF诱导获能精子的顶体反应,但对未获能或无钙培养基中的精子无影响[23]。虽然我们目前的研究表明,Lyso-PAF并不能诱导顶体反应,但有一项研究显示,Lyso-PAF处理对顶体反应的诱导和人精子的穿透率有积极作用[24]。
我们研究的初步结果表明,PAF处理精子90分钟对精子活力没有任何影响。然而,之前的研究结果却显示PAF浓度与精子活力之间存在正相关关系。对恒河猴精子的研究显示内源性PAF浓度与精子活力和进展相关[6]。据报道,PAF治疗可增加运动精子的数量和冷冻精子的运动参数[25,27]。PAF治疗4小时后,25份精子样本中有16份活力增加,并且过夜治疗后增强了所有精子样本的运动性[22]。Grassi G等人报道外源性使用浓度为0.1M的PAF持续治疗60分钟可显着提高精子活力。此外,用PAF受体拮抗剂CV-3988治疗24小时表现出对精子活力的抑制和可逆作用[26]。因此,PAF对精子活力的调节作用可能与潜伏期的持续时间有关。
PAF受体位于人类精子的中段和近端头部[29]。PAF与人精子表面互补受体的结合相互作用诱导肌醇三磷酸的形成,通过刺激细胞内钙的释放,促进外源钙通过质膜钙通道的流入,增加细胞内钙浓度[30,31]。细胞内钙浓度升高可诱导膜间肌动蛋白网络解聚,从而激活与顶体反应相关的磷脂酶。PAF拮抗剂和PAF受体抗体可抑制精子活力、顶体反应率和精子穿透卵母细胞的能力[23,35]。酪氨酸磷酸化涉及精子功能的各个方面,包括顶体反应和精子-透明带结合[11]。获能过程中,酪氨酸磷酸化蛋白从非获能性精子的尾部移位到获能或透明带暴露的精子的顶体区域[11]。本研究结果表明,PAF诱导的人精子蛋白酪氨酸磷酸化可能与顶体反应率的正向影响有关。
细胞外信号调节激酶(ERK)属于丝裂原活化蛋白激酶(MAPK)信号传导途径家族。原型ERK级联主要由Raf(MAPK激酶激酶)、MEK1/2(MAPK激酶)和ERK(ERK1和ERK2)组成[36]。Raf启动MEK的磷酸化,其反过来使ERK1和ERK2磷酸化(ERK1和ERK2也称为p44和p42 MAPK)。我们的研究结果证实,PAF通过促进ERK1和ERK2的表达激活人精子中的ERK信号通路。
值得注意的是,ERK拮抗剂U0126治疗降低了PAF诱导的顶体反应,而对自发顶体反应没有影响。结果表明,PAF可通过激活ERK通路提高顶体反应率。
ERKs定位于人精子顶体后区,并在顶体反应时重新分布于赤道区[37]。ERK级联在精子顶体反应调控中的作用尚不明确。先前的研究表明,ERK通路参与了精子活力的提高、头状位的形成以及PKC介导的顶体反应[37-39]。在体外获能期间,ERK1和ERK2以时间依赖性方式被酪氨酸磷酸化[37]。MAPK抑制剂PD098059能显著抑制透明带、钙离子团A23187和孕酮诱导的顶体反应[13]。另一方面,有研究报道ERK活化与精子受精潜能降低有关。据报道,ERK1表达的增加与精子质量低下有关。ERK的下调可增强人类精子中透明带诱导的顶体反应[13]。综上所述,ERK对PAF诱导的顶体反应的确切作用有待进一步研究。
PAF明显存在于人类精子中。PAF活化酶(PAF-乙酰转移酶)和失活酶(PAF-乙酰水解酶)在哺乳动物精子和精液中都有丰富的表达[41]。这提示附睾液中PAF-乙酰水解酶的存在可使精子中的PAF失活,防止其过早获能。当射精的精子通过女性生殖道时,PAF被激活并随后启动生物活动,引起精子活力、顶体反应和精子与卵母细胞的相互作用增强[23,27,44]。
总之,PAF诱导精子顶体反应和酪氨酸磷酸化。PAF对精子顶体反应的积极作用可能是通过ERK信号介导的。我们的研究结果进一步验证了PAF对人类精子受精能力的潜在作用。
参考文献
[1]Florman HM,Jungnickel MK,Sutton KA.Regulating the acrosome reaction[J].The International journal of developmental biology,2008,52:503-10.
[2]Hirohashi N,Yanagimachi R.Sperm acrosome reaction:its site and role in fertilization[J].Biology of reproduction,2018,99:127-33.
[3]Hanahan DJ.Platelet activating factor:a biologically active phosphoglyceride[J].Annual review of biochemistry,1986,55:483-509.
[4]Prescott SM,Zimmerman GA,Stafforini DM,et al.Platelet-activating factor and related lipid mediators[J].Annual review of biochemistry,2000,69:419-45.
[5]Minhas BS,Kumar R,Ricker DD,et al.The presence of platelet-activating factor-like activity in human spermatozoa[J].Fertility and sterility,1991,55:372-6.
[6]Roudebush WE,Gerald MS,Cano JA,et al.Relationship between platelet-activating factor concentration in rhesus monkey(Macaca mulatta)spermatozoa and sperm motility[J].American journal of primatology,2002,56:1-7.
[7]Minhas BS:Platelet-activating factor treatment of human spermatozoa enhances fertilization potential[J].American journal of obstetrics and gynecology,1993,168:1314-7.
[8]Lecewicz M,Kordan W,Kaminski S,et al.Effects of the platelet-activating factor(PAF)supplementation on ATP content of cryopreserved bull spermatozoa(AI)[J].Polish journal of veterinary sciences,2017,20:421-3.
[9]Kyriakis JM.Making the connection:coupling of stress-activated ERK/MAPK(extracellular-signal-regulated kinase/mitogen-activated protein kinase)core signalling modules to extracellular stimuli and biological responses[J].Biochemical Society symposium,1999,64:29-48.
[10]Cocchia N,Pasolini MP,Mancini R,et al.Effect of sod(superoxide dismutase)protein supplementation in semen extenders on motility,viability,acrosome status and ERK(extracellular signal-regulated kinase)protein phosphorylation of chilled stallion spermatozoa[J].Theriogenology,2011,75:1201-10.
[11]Naz RK,Rajesh PB.Role of tyrosine phosphorylation in sperm capacitation/acrosome reaction[J].Reproductive biology and endocrinology:RB&E,2004,2:75.
[12]De Lamirande E,Gagnon C:The extracellular signal-regulated kinase(ERK)pathway is involved in human sperm function and modulated by the superoxide anion[J].Molecular human reproduction,2002,8:124-35.
[13]Du Plessis SS,Page C,Franken DR.The zona pellucida-induced acrosome reaction of human spermatozoa involves extracellular signal-regulated kinase activation[J].Andrologia,2001,33:337-42.
[14]Chiu PC,Ho PC,Ng EH,et al.Comparative study of the biological activity of spermatozoa-zona pellucida binding inhibitory factors from human follicular fluid on various sperm function parameters[J].Molecular reproduction and development,2002,61:205-12.
[15]Zimmerman GA,McIntyre TM,Prescott SM,et al.The platelet-activating factor signaling system and its regulators in syndromes of inflammation and thrombosis[J].Critical care medicine,2002,30:294-301.
[16]Ishii S,Shimizu T.Platelet-activating factor(PAF)receptor and genetically engineered PAF receptor mutant mice[J].Progress in lipid research,2000,39:41-82.
[17]Roudebush WE,Toledo AA,Kort HI,et al.Platelet-activating factor significantly enhances intrauterine insemination pregnancy rates in non-male factor infertility[J].Fertility and sterility,2004,82:52-6.
[18]Toledo AA,Mitchell-Leef D,Elsner CW,et al.Fertilization potential of human sperm is correlated with endogenous platelet-activating factor content[J].Journal of assisted reproduction and genetics,2003,20:192-5.
[19]Kumar S,Sharma A.Platelet activating factor improves the in vitro penetration of zona free hamster eggs by buffalo(Bubalus bubalis)spermatozoa[J].Theriogenology,2005,63:1564-72.
[20]Roudebush WE,Fukuda AI,Minhas BS.Enhanced embryo development of rabbit oocytes fertilized in vitro with platelet activating factor(PAF)-treated spermatozoa[J].Journal of assisted reproduction and genetics,1993,10:91-4.
[21]Kordan W,Lecewicz M,Tobolski J.Effect of platelet-activating factor on motility,plasmalemma integrity,the process of capacitation and acrosome reaction of fresh and cryopreserved boar spermatozoa[J].Polish journal of veterinary sciences,2009,12:175-81.
[22]Krausz C,Gervasi G,Forti G,et al.Effect of platelet-activating factor on motility and acrosome reaction of human spermatozoa[J].Human reproduction,1994,9:471-6.
[23]Angle MJ,Tom R,Jarvi K,et al.Effect of platelet-activating factor(PAF)on human spermatozoa-oocyte interactions[J].Journal of reproduction and fertility,1993,98:541-8.
[24]Lachapelle MH,Bouzayen R,Langlais J,et al.Effect of lysoplatelet-activating factor on human sperm fertilizing ability[J].Fertility and sterility,1993,59:863-8.
[25]Briton-Jones C,Yeung QS,Tjer GC,et al.The effects of follicular fluid and platelet-activating factor on motion characteristics of poor-quality cryopreserved human sperm[J].Journal of assisted reproduction and genetics,2001,18:165-70.
[26]Sengoku K,Tamate K,Takaoka Y,et al.Effects of platelet activating factor on human sperm function in vitro[J].Human reproduction,1993,8:1443-7.
[27]Hellstrom WJ,Wang R,Sikka SC.Platelet-activating factor stimulates motion parameters of cryopreserved human sperm[J].Fertility and sterility,1991,56:768-70.
[28]Grassi G,Cappello N,Gheorghe MF,,et al.Exogenous platelet-activating factor improves the motility of human spermatozoa evaluated with C.A.S.A.:optimal concentration and incubation time[J].Journal of endocrinological investigation,2010,33:684-90.
[29]Roudebush WE,Wild MD,Maguire EH.Expression of the platelet-activating factor receptor in human spermatozoa:differences in messenger ribonucleic acid content and protein distribution between normal and abnormal spermatozoa[J].Fertility and sterility,2000,73:967-71.
[30]Shukla SD.Platelet activating factor-stimulated formation of inositol triphosphate in platelets and its regulation by various agents including Ca2+,indomethacin,CV-3988,and forskolin[J]..Archives of biochemistry and biophysics,1985,240:674-81.
[31]Logan JE,Roudebush WE.Platelet-activating factor increases intracellular calcium levels in preimplantation stage embryos[J]..Early pregnancy,2000,4:30-8.
[32]Bragado MJ,Gil MC,Garcia-Marin LJ.Platelet-activating factor in Iberian pig spermatozoa:receptor expression and role as enhancer of the calcium-induced acrosome reaction[J].Reproduction in domestic animals=Zuchthygiene,2011,46:943-9.
[33]Guzman RE,Bolanos P,Delgado A,et al.Depolymerisation and rearrangement of actin filaments during exocytosis in rat peritoneal mast cells:involvement of ryanodine-sensitive calcium stores[J].Pflugers Archiv:European journal of physiology,2007,454:131-41.
[34]Fedorov P,Dzyuba B,Fedorova G,et al.Quantification of adenosine triphosphate,adenosine diphosphate,and creatine phosphate in sterlet spermatozoa during maturation[J].Journal of animal science,2015,93:5214-21.
[35]Kuzan FB,Geissler FT,Henderson WR,et al.Role of spermatozoal platelet-activating factor in fertilization[J].Prostaglandins,1990,39:61-74.
[36]Kolch W.Meaningful relationships:the regulation of the Ras/Raf/MEK/ERK pathway by protein interactions[J].The Biochemical journal,2000,351(2):289-305.
[37]Luconi M,Barni T,Vannelli GB,et al.Extracellular signal-regulated kinases modulate capacitation of human spermatozoa[J].Biology of reproduction,1998,58:1476-89.
[38]Thundathil J,de Lamirande E,Gagnon C,et al.Different signal transduction pathways are involved during human sperm capacitation induced by biological and pharmacological agents[J].Molecular human reproduction,2002,8:811-6.
[39]Almog T,Lazar S,Reiss N.Identification of extracellular signal-regulated kinase 1/2 and p38 MAPK as regulators of human sperm motility and acrosome reaction and as predictors of poor spermatozoan quality[J].The Journal of biological chemistry,2008,283:14479-89.
[40]Chiu PC,Wong BS,Lee CL,et al.Zona pellucida-induced acrosome reaction in human spermatozoa is potentiated by glycodelin-A via down-regulation of extracellular signal-regulated kinases and up-regulation of zona pellucida-induced calcium influx[J].Human reproduction,2010,25:2721-33.
[41]Angle MJ,Tom R,Khoo D,et al.Platelet-activating factor in sperm from fertile and subfertile men[J].Fertility and sterility,1991,56:314-8.
[42]Tjoelker LW,Eberhardt C,Unger J,et al.Plasma platelet-activating factor acetylhydrolase is a secreted phospholipase A2 with a catalytic triad[J].The Journal of biological chemistry,1995,270:25481-7.
[43]Muguruma K,Johnston JM.Metabolism of platelet-activating factor in rat epididymal spermatozoa[J].Biology of reproduction,1997,56:529-36.
[44]Roudebush WE,Minhas BS,Ricker DD,et al.Platelet activating factor enhances in vitro fertilization of rabbit oocytes[J].American journal of obstetrics and gynecology,1990,163:1670-3.
关注SCI论文创作发表,寻求SCI论文修改润色、SCI论文代发表等服务支撑,请锁定SCI论文网! 文章出自SCI论文网转载请注明出处:https://www.lunwensci.com/yixuelunwen/17444.html